
Для термина «волокно» см. также другие значения.
Не́рвные воло́кна
- длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие[1][2].
миелиновые
и
безмиелиновые
- Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого
осевым цилиндром
(аксоном), и, в случае миелиновых волокон, окружающей его оболочкой. В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый (мякотный) тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый (безмякотный) тип.
Покрытые оболочкой сплетения пучков нервных волокон образуют нервы.
Классификация Эрлангера-Гассера[ | ]
Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
| Aα | Афферентные — мышечные веретёна, сухожильные органы; эфферентные — скелетные мышцы | 10-20 | 60-120 | + |
| Aβ | Афферентные — тактильное чувство; коллатерали Aα волокон к интрафузальным мышечным волокнам | 7-15 | 40-90 | + |
| Aγ | Эфферентные — мышечные веретёна | 4-8 | 15-30 | + |
| Aδ | Афферентные — температура, быстрое проведение боли | 3-5 | 5-25 | + |
| B | Симпатические, преганглионарные; постганглионарные волокна цилиарного ганглия | 1-3 | 3-15 | прерывистая |
| C | Симпатические, постганглионарные; афферентные — медленное проведение боли | 0,3-1 | 0,5-2 | — |
Строение безмиелинового нервного волокна
- представляет собой цепь леммоцитов, в которую вдавлено от 5 до 20 осевых цилиндров. Каждый осевой цилиндр прогибает цитолемму леммоцита и вдавливается в его цитоплазму. Сближенные участки цитолеммы образуют мезаксон
(дословно – брыжейка аксона).
На поперечном срезе безмиелинового нервного волокна определяются:
v в центре волокна – ядро леммоцита
v по периферии – поперечные сечения осевых цилиндров, как бы подвешенные на мезаксонах.
v по самой периферии волокна – базальная пластинка из соединительной ткани. Безмиелиновые нервные волокна относятся к волнам кабельного типа, они тонкие (5-7- мкм) и проводят нервные импульсы очень медленно (1-2- м/сек).
Строение миелинового нервного волокна.
Ø Осевой цилиндр один, погружен в центральную часть цепи леммоцитов.
Ø Мезаксон длинный, закручен вокруг осевого цилиндра, образуя миелиновый слой.
Ø Цитоплазма и ядро леммоцитта сдвигаются на периферию и образуют неврилемму нервного волокна.
Ø Снаружи расположена базальная пластинка.
узловые перехваты (перехваты Ранвье)
- Участок между 2 перехватами называется
межузловым сегментом
- В каждом межузловом сегменте отчётливо видны насечки миелина – прозрачные участки, в которых содержится цитоплазма леммоцита между витками мезаксона.
Диаметр миелиновых нервных волокон 12-20 мкм, скорость проведения нервного импульса – большая (1о-120 м/с).
Высокая скорость проведения импульсов объясняется сальтаторным способом передачи – скачками от 1 перехвата Ранвье к другому.
Нерв – комплексное образование, состоящее из:
1) миелиновых и безмиелиновых нервных волокон
2) рыхлой волокнистой соединительной ткани, образующей оболочки нерва:
эндоневрий
- окружает отдельные нервные волокна
периневрий
- окружает пучки нервных волокон
эпиневрий
- окружает нервный ствол.
В оболочках проходят кровеносные сосуды, обеспечивающие трофику нервов.
НЕРВНЫЕ ОКОНЧАНИЯ
- это концевые разветвления отростков нервных клеток, в которых нервный импульс или генерируется или передается на другую клетку.
3 группы:
1) Эффекторные (двигательные)
2) Рецепторные (чувствительные)
3) Межнейронные синапсы.
Если осевой цилиндр нервного волокна является дендритом чувствительного нейрона, то его концевой аппарат образует рецептор. Если осевой цилиндр является аксоном нервной клетки, его концевой аппарат образует эффекторное или синаптическое окончание.
ДВИГАТЕЛЬНОЕ НЕРВНОЕ ОКОНЧАНИЕ
3 основные части
1) нервный полюс
2) синаптическая щель
3) мышечный полюс
Нервный полюс
В нервном полюсе содержатся следующие элементы:
- скопление митохондрий с продольными кристами
- синаптические пузырьки с медиатором (а/х).
- пресинаптическая мембрана.
Синаптическая щель
- пространство в 50 нм между пре- и постсинаптическими мембранами, в котором находится фермент а/хэстераза.
Мышечный полюс включает в себя:
- постсинатпическую мембрану – участок плазмолеммы миосимпласта, содержащий рецепторы к а/х
- участок саркоплазмы миосимпласта, в котором отсутствуют миофибриллы и содержится скопление ядер и саркосом.
Прохождение нервного импульса по нервному окончанию вызывает выделение а/х из синаптических пузырьков через пресинаптическую мембрану. А/х вызывает возбуждение рецепторов постсинаптической мембраны и появление волны деполяризации, которая распространяется по плазмолемме миосимпласта, а затем и по Т-канальцам. Затем деполяризация передается на цистерны саркоплазматической сети, вызывая выход ионов кальция и сокращение мышечного волокна. Наличие а/хэстеразы в синаптической щели обусловливает дискретное действие а/х на рецепторы постсинаптической мембраны, и кратковременное сокращение мышечного волокна.
РЕЦЕПТОРЫ, КЛАССИФИКАЦИЯ, По локализации
- интерорецепторы – рецепторы внутренних органов
- экстерорецепторы – рецепторы , воспринимающие раздражения из внешней среды
- поприорецепторы – воспринимают информацию от опрно-двигательного аппарата.
По специфичности восприятия
- хеморец
- механо-
- баро –
- термо-
- ноци –
Классификация по Ллойду[ | ]
Классифицирует только
афферентные нейроны.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
| Ia | Мышечные веретёна | 18-22 | 90-120 | + |
| Ib | Сухожильные рецепторы | 15-18 | 60-90 | — |
| II | Механорецепторы кожи, вторичные мышечные веретёна | 7-15 | 40-90 | + |
| III | Рецепторы связок | 1-5 | 3-25 | прерывистая |
| IV | Болевые рецепторы, рецепторы соединительной ткани | 0,1-1 | 0,5-2 | — |
Физиология нервных волокон и нервов
2.1.1. Структура нервного волокна
безмиелиновых волокон
образуют шванновские клетки (леммоциты).
При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окружает каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон).
миелиновых волокон
образуют в периферической нервной системе также шванновские клетки, а в ЦНС — олигодендроциты. В отличие от безмиелиновых волокон в миелиновых волокнах мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, образуя слой миелина толщиной от долей мкм до 10 мкм (липидный футляр) вокруг осевого цилиндра. Миелиновая оболочка через равные участки (0,5— 2,0 мм) прерывается, образуя свободные от миелина небольшие участки — узловые перехваты Ранвье. Протяженность перехватов в волокнах периферической нервной системы находится в пределах 0,25—1,0 мкм, в волокнах ЦНС их длина может достигать 14 мкм. Участки волокон между узловыми перехватами называются межузловыми сегментами, они образованы слоем миелина. Основную часть миелина (78 % сухой массы) составляют липиды, в них на долю фосфолипидов приходится 42 %, цереброзидов — 28 %, холестерина — 25 %. Несмотря на метаболическую инертность миелина (особенно по обновлению холестерина), поддержание целостности его структуры требует затраты энергии и нарушение снабжения олигодендроцитов кислородом и питательными веществами быстро вызывает деструкцию миелина. Одной из основных функций глиальной оболочки нервных волокон является изолирующая функция, способствующая лучшему проведению биопотенциалов по отросткам нейронов.
2.1.2.Классификация нервных волокон
Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С (табл.3).
Волокна типа А и В являются миелиновыми, типа С — безмиелиновыми. Волокна А делят на 4 подгруппы: α, β, γ, δ. В периферической нервной системе к волокнам Аα относятся афферентные волокна от механорецепторов кожи, мышечных и сухожильных рецепторов, а также эфферентные волокна к скелетным мышцам. К Аβ принадлежат афферентные волокна от кожных рецепторов прикосновения и давления, от части мышечных и висцеральных рецепторов. Аγ представляют собой эфферентные волокна, через которые регулируется активность мышечных рецепторов. К Аδ относят афферентные волокна от части тактильных, температурных и болевых, а также суставных рецепторов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы. К волокнам типа С относят постганглионарные волокна вегетативной нервной системы, афферентные волокна от некоторых болевых (вторичная боль), тепловых и висцеральных рецепторов.
Таблица 3. Типы волокон в нервах млекопитающих (по Эрлангеру—Гассеру)
| Тип волокон | Диаметр волокна, мкм | Скорость проведения возбуждения, м/с | Длительность абсолютного рефракторного периода, мс |
| Аa | 12-20 | 70-120 | 0,4-1,0 |
| Аb | 5-12 | 30-70 | |
| Аg | 3-6 | 15-30 | |
| Аd | 2-5 | 12-30 | |
| В | 1-3 | 5-12 | 1,2 |
| С | 0,3-1,3 | 0,5-2,3 |
Из данных, представленных в табл.3, видно, что средний диаметр каждого типа волокна снижается от типа А до С (каждый примерно в 2 раза по отношению к предыдущему).
Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аα до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон Аα (наибольшая возбудимость) к волокнам С (наименьшая возбудимость).
Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон Аα. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к местным анестетикам — волокна С.
Нервные волокна имеют две основные функции — проведение возбуждения и транспорт веществ, обеспечивающих трофическую функцию.
2.1.3.Механизм проведения возбуждения по нервному волокну
локальными
(местными), распространяющимися с декрементом (затуханием) на расстояние, не превышающее 1—2 мм, и
импульсными
(ПД), распространяющимися без декремента по всей длине волокна — на несколько десятков сантиметров, например от мотонейронов спинною мозга по всей длине нервного волокна до мышечных волокон конечностей с учетом длины самой конечности.
Распространение локальных потенциалов.
электротонической.
Электротоническое распространение возбуждения — физический механизм, оно характерно для тех фрагментов мембран возбудимых клеток, где нет потенциалзависимых ионных каналов. Такими участками являются, например, большая часть мембраны дендритов нервных клеток, межперехватные промежутки в миелиновых нервных волокнах. Если местный потенциал (рецепторный или ВПСП), распространяясь электротонически, достигает участков мембраны, способных генерировать ПД (аксонный холмик, перехваты Ранвье, часть мембраны дендритов и, возможно, сомы), но его амплитуда при этом не достигнет критического уровня деполяризации, то такой потенциал называют препотенциалом. В его возникновении и распространении частично участвуют потенциалзависимые ионные каналы, однако при этом нет регенеративной (самоусиливающейся) деполяризации, характерной для ПД. Поэтому распространение такого потенциала происходит с затуханием амплитуды. Если локальный потенциал достигает участков мембраны, способных генерировать ПД, и его амплитуда выходит на критический уровень деполяризации, формируется ПД, который распространяется по всей длине нервного волокна без затухания.
Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна — сопротивления и емкости его мембраны, сопротивления цитоплазмы. Электротоническое проведение в нервном волокне улучшается при увеличении его диаметра, что связано с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, увеличивающей сопротивление мембраны (до 105 Ом/см2) и уменьшающей ее емкость (до 0,005 мкФ/см2).
Эффективность электротонического проведения характеризует постоянная длины мембраны (λm).
Это расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37 % от исходной величины. Постоянная длины для локальных потенциалов реально не превышает 1 мм, и их амплитуда затухает на расстоянии 1—2 мм от места возникновения.
Для передачи возбуждения на большие расстояния необходимо формирование ПД. В его распространении, кроме электротонического механизма, существенная роль принадлежит механизму регенеративной деполяризации, позволяющей сохранить амплитуду ПД на всем пути его следования.
Проведение потенциала действия., Непрерывное распространение ПД
Поэтому ПД, распространяясь на этом расстоянии как электротонический потенциал и сохранив как минимум 37 % своей амплитуды, способен деполяризовать мембрану до критического уровня и генерировать на всем протяжении новые ПД (рис.5).
При этом на этапе электротонического распространения нервного импульса ионы движутся вдоль волокна между деполяризованным и поляризованным участками, обеспечивая проведение возбуждения в соседние участки волокна. Реально при неповрежденном нервном волокне этап чисто электротонического распространения ПД (вдоль мембраны) предельно мал, так как потенциалзависимые каналы имеются в непосредственной близости друг от друга и, естественно, — от возникшего потенциала действия и наблюдается только до достижения деполяризации, равной 50 % Екр. Далее включается перемещение ионов в клетку (нервное волокно) и из клетки за счет активации ионных каналов.
При формировании нового ПД в соседнем участке в фазе деполяризации возникает мощный ток ионов натрия в клетку вследствие активации натриевых каналов, приводящий к регенеративной (самоусиливающейся) деполяризации. Этот ток обеспечивает формирование нового ПД той же амплитуды, представляющий собой, как обычно, сумму двух величин — мембранного потенциала покоя и инверсии. В связи с этим проведение ПД осуществляется без декремента (без снижения амплитуды).
Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый (при поступлении к нему электротонического потенциала), а затем как раздражающий (после формирования в нем нового ПД).
Сальтаторный ( прерывистый, скачкообразный )
нервного импульса осуществляется в миелиновых волокнах (типы А и В), для которых характерна концентрация потенциалзависимых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 12 000 на 1 мкм2, что примерно в 100 раз выше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт (межузловых сегментов), обладающих хорошими изолирующими свойствами, потенциалзависимых каналов почти нет, и мембрана осевого цилиндра там практически невозбудима. В этих условиях ПД, возникший в одном перехвате Ранвье, электротонически (вдоль волокна, без участия ионных каналов) распространяется до соседнего перехвата, деполяризуя там мембрану до критического уровня, что приводит к возникновению нового ПД, т.е. возбуждение проводится скачкообразно (рис. 6).
Постоянная длина мембраны миелинового волокна достигает 5 мм. Это значит, что ПД, распространяясь электротонически на этом расстоянии, сохраняет 37 % своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мВ).
Поэтому в случае повреждения ближайших на пути следования перехватов Ранвье потенциал действия может электротонически возбудить 2—4-й и даже 5-й перехваты.
Во-первых,
оно более экономично в энергетическом плане, т.к. возбуждаются только перехваты Ранвье, площадь которых менее 1 % мембраны, и, следовательно, надо меньше энергии для восстановления трансмембранных градиентов Na+ и К+, уменьшающихся в процессе формирования ПД.
Во-вторых,
возбуждение проводится с большей скоростью (см. табл.3), чем в безмиеликовых волокнах, так как возникший ПД на протяжении миелиновых муфт распространяется электротонически, что в 107 раз быстрее, чем скорость непрерывного проведения ПД в безмиелиновом волокне.
2.1.4. Проведение возбуждения в нервных стволах
В периферической нервной системе волокна объединены с помощью соединительнотканных оболочек в нервные стволы (нервы).
В одном нерве могут быть тысячи нервных волокон: например, в срединном и мышечно-кожном нервах имеется 27—37 тыс. нервных волокон. Волокна в нервах могут быть миелиновыми и безмиелиновыми, афферентными и эфферентными. В естественных условиях каждое волокно нерва возбуждается от своего источника (например, эфферентное — от аксонного холмика, афферентное — от рецептора), и ПД в них проводятся асинхронно. Кроме того, чувствительные и двигательные волокна проводят импульсы в противоположных направлениях. Суммарная электрическая активность нерва создается электрической активностью составляющих его волокон и зависит от числа возбужденных волокон, степени шунтирования местных токов невозбужденными волокнами, синхронности проведения ПД в волокнах. В связи с этим анализ суммарной электрической активности нерва (нейрограммы) представляет трудную задачу.
В лабораторных условиях при монополярном отведении, когда один электрод расположен на неповрежденном участке нерва, а второй — на поврежденном (деполяризованном) участке, можно зафиксировать суммарный монофазный ПД нерва и его дисперсию (расслоение) во времени. Если отводящий электрод расположен близко (до 3 мм) к раздражающему, через который подают сильный одиночный стимул, то регистрируется суммарный ПД нерва, напоминающий по форме ПД отдельного нервного волокна, но растянутый по времени. Суммарный ПД нерва в отличие от ПД отдельного волокна не подчиняется закону «все или ничего». Это означает, что при увеличении силы раздражения увеличивается число возбужденных нервных волокон: в возбуждение вовлекаются, кроме Аα-волокон, менее возбудимые Аβ-, Аγ-, Аδ-, В-волокна и, наконец, наименее возбудимые С-волокна (закон силовых отношений — увеличение ответной реакции с увеличением силы раздражения).
Если отводящий электрод расположен на достаточном удалении от раздражающего электрода (до 80—100 мм), то фиксируется расслоение суммарного ПД нерва на несколько пиков соответственно типам нервных волокон. Это связано с неодинаковой скоростью проведения ПД в разных волокнах нерва: сначала до места регистрации доходят нервные импульсы по быстропроводящим Аα-волокнам, через некоторое время по Аβ, затем по Аγ и т.д. Позже всего до места регистрации доходят ПД по С-волокнам.
Если отведение биполярное и оба отводящих электрода расположены на неповрежденных участках нерва и недалеко от раздражающего электрода (чтобы избежать дисперсии потенциала, то при сильном одиночном стимуле фиксируется двухфазный суммарный потенциал. Возникновение этих фаз связано с тем, что, когда волна возбуждения находится под первым (ближайшим к месту раздражения) отводящим электродом, этот участок становится электроотрицательным по отношению к покоящемуся участку под вторым отводящим электродом и луч осциллографа отклоняется вверх. Когда же волна возбуждения доходит до второго электрода, а под первым электродом мембраны волокон уже реполяризованы, то луч осциллографа отклоняется в противоположную сторону — вниз.
2.1.5. Законы проведения возбуждения по нервным волокнам
− Закон двустороннего проведения возбуждения.
− Закон изолированного проведения возбуждения .
− Закон физиологической непрерывности нерва.
Возможность функционального блока проведения возбуждения возможна при морфологической целостности волокон. Непроводимость наступает при воздействиях, нарушающих генерацию нервного импульса. Так, чрезмерное охлаждение или согревание, прекращение кровоснабжения, различные химические агенты, в частности местные обезболивающие — новокаин, кокаин, дикаин, прекращают проведение по нерву. Н.Е.Введенский (1901) показал, что при действии различных факторов на нерв (кокаина, хлороформа, фенола, хлористого калия, сильного фарадического тока) в нем сначала возникает трансформация ритма проводимого возбуждения (блокируется проведение высокочастотных потенциалов действия, и проводятся только низкочастотные ПД), а в дальнейшем может возникать полный блок проведения нервных импульсов — участок парабиоза. В этом участке возникает длительная деполяризация мембраны волокон, которая в результате закрытия инактивационных h-ворот в натриевых каналах сначала затрудняет генерацию ПД (уменьшается его амплитуда, увеличивается длительность, затягивается фаза абсолютной рефрактерности), а в дальнейшем, если инактивация натриевых каналов превысит 50 %, приводит к полной невозбудимости этого участка нервного волокна. Для возникновения блока в проведении возбуждения протяженность парабиотического участка должна превысить постоянную длину мембраны (λm), иначе ПД может распространиться через этот участок электротонически. Нарушение физиологической непрерывности нервных волокон возникает при действии анестетиков, электрического тока, при гипоксии, воспалении, охлаждении. После прекращения действия этих факторов проведение возбуждения по волокнам нерва восстанавливается. Однако, при углублении и усилении действия вызвавшего парабиоз агента обратимые изменения могут переходить в необратимое нарушение жизнедеятельности — смерть.
2.1.6. Особенности проведения возбуждения в нервных волокнах
− Большая скорость проведения возбуждения.
− Малая утомляемость нервного волокна.
Развернуть
Открыть в широком формате
Миелинизация нервных волокон[ | ]
При формировании безмиелинового нервного волокна осевой цилиндр (отросток нейрона) погружается в тяж из леммоцитов, цитолеммы которых прогибаются и плотно охватывают осевой цилиндр в виде муфты, края которой смыкаются над ним, образуя дупликатуру клеточной мембраны — мезаксон
— Соседние леммоциты входящие в состав сплошного глиального тяжа своими цитолеммами образуют простые контакты. Безмиелиновые нервные волокна имеют слабую изоляцию, допускающую переход нервного импульса с одного волокна на другое, как в области мезаксона, так и в области межлеммоцитарных контактов.
шванновской оболочкой
или
неврилеммой
— Внутренний слой, состоящий из витков мезаксона, называется миелиновым или миелиновой оболочкой. Следствием того, что миелинизация происходит в процессе роста как отростков нейронов, так и самих леммоцитов, является постепенное увеличение количества витков и размеров мезаксона, то есть каждый последующий виток шире предыдущего. Следовательно, последний виток, содержащий цитоплазму с ядром леммоцита является самым широким. Толщина миелина по длине волокна неоднородна, а в местах контактов соседних леммоцитов слоистая структура исчезает и контактируют лишь наружные слои, содержащие цитоплазму и ядро. Места их контактов называются
узловыми перехватами
перехватами Ранвье
, возникающими вследствие отсутствия здесь миелина и истончения волокна.
В ЦНС миелинизация нервного волокна происходит за счет обхвата осевых цилиндров отростками олигодендроцитов.
насечки миелина
насечки Шмидта-Лантермана
, которые соответствуют небольшим прослойкам цитоплазмы леммоцита.
аксолеммой
— Она обеспечивает проведение нервного импульса, который представляет собой волну деполяризации аксолеммы. Если осевой цилиндр представлен нейритом, то в нём отсутствуют гранулы базофильного вещества.
Гистология.RU , Материал взят с сайта www.hystology.ru
Отростки нервных клеток в совокупности с покрывающими их клетками нейроглии образуют нервные волокна. Расположенные в них отростки нервных клеток (дендриты или нейриты) называют осевыми цилиндрами, а покрывающие их клетки олигодендроглпи — нейролеммоцитами (леммоцитами, шванновскими клетками).
В соответствии с составом нервных волокон и морфологическими особенностями их строения различают миелиновые и безмиелиновые нервные волокна.
Безмиелиновые (безмякотные) нервные волокна
Рис. 159. Безмиелиновые нервные волокна (по Кахалу).
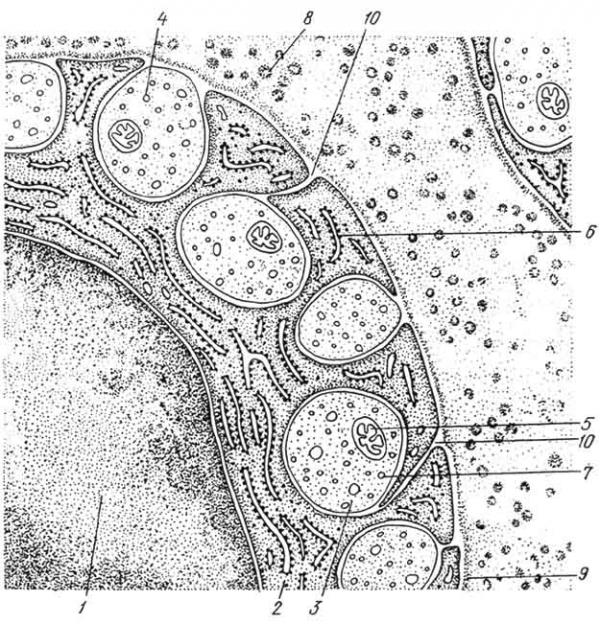
Рис. 160. Схема строения безмиелинового нервного волокна:
1 — ядро и 2 — цитоплазма леммоцита; 3 — 4 — аксоны нервных клеток, погруженные в цитоплазму леммоцита (3 — полностью, 4 — частично); 5 — митохондрии; 6 — шероховатая эноплазматическая сет; 7 — микротрубочки; 8 — коллагеновые волокна эндоневрия; 9 — базальная мембрана; 10 — мезаксон
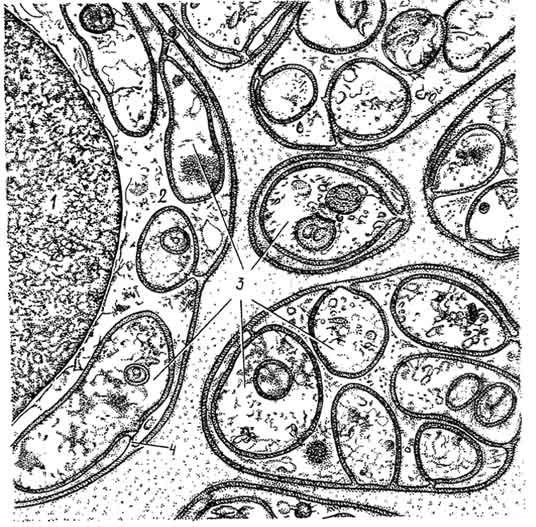
Рис. 161. Электронная микрофотография поперечного среза безмиелиновых нервных волокон брыжейки крысы:
1 — ядро леммоцита; 2 — цитоплазма леммоцита; 3 — осевые цилиндры; 4 — мезаксон (по Фаусетту).
клетки прогибается, образуя более или менее глубокие складки, содержащие отростки нервных клеток — осевые цилиндры. Сближенные в области складки участки плазмолеммы леммоцита формируют сдвоенную мембрану — мезаксон, на котором подвешен осевой цилиндр. При световой микроскопии плазмолемма леммоцита и мезаксоны не выявляются, что создает впечатление погружения осевых цилиндров непосредственно в цитоплазму глиальных клеток (рис. 159, 160, 161).
Миелиновые (мякотные) нервные волокна
Рис. 162. Миелиновые нервные волокна из седалищного нерва лягушки, обработанные осмиевой кислотой:
1 — осевой цилиндр; 2 — леммоцит; 3 — миелиновая оболочка; 4 — соединительная ткань; 5 — насечки; 6 — перехват.
два слоя: внутренний — миелиновый, более толстый и наружный — тонкий, содержащий цитоплазму и ядра леммоцитов.
На границе двух леммоцитов оболочка миелинового волокна истончается, образуется сужение волокна — узловой перехват (перехват Ранвье).
Участок нервного волокна между двумя узловыми перехватами называется межузловым сегментом. Его оболочка соответствует одному леммоциту (рис. 162).
В процессе развития миелинового нервного волокна осевой цилиндр, погружаясь в цитоплазму леммоцита, вовлекает с собой его плазмолемму. Здесь, как и в безмиелиновом волокне, формируется мезаксон, который, последовательно удлиняясь и концентрически наслаиваясь на осевой цилиндр, образует зону плотно расположенных мембран — миели-новую оболочку волокна (рис. 163, 164).
При электронной микроскопии видно, что мембраны мезаксона образуют чередующиеся светлые линии (8 — 12 нм), соответствующие их липидным слоям, и темные тонкие, образованные белковыми молекулами (рис. 165).
Наружный слой оболочки миелинового волокна — неврилемма (шванновская оболочка) — соответствует оттесненной на периферию цитоплазме леммоцитов с их ядрами. Плотно расположенные витки мезаксона в соответствующих участках волокна, отклоняясь от параллельной осевому цилиндру ориентации, рассредоточиваются и формируют косо ориентированные зоны оболочки, более богатые цитоплазмой. При осмировании волокна они выделяются как светлые линии — насечки миелина.
В области перехвата слои мембран миелиновой оболочки контактируют с осевым цилиндром волокна. В соответствии с последовательностью роста осевого цилиндра и формирования слоев мезаксона более глубокие слои последнего короче поверхностных и располагаются дальше от перехвата.
Смежные леммоциты оболочки волокна взаимодействуют друг € другом системой более или менее выраженных пальцевидных отростков, которые, переплетаясь, формируют их контакты и на срезах наблюдаются в различных сечениях. Миелиновое волокно с поверхности покрыто базальной мембраной, связанной с тяжами коллагеновых волокон окружающей соединительной ткани.
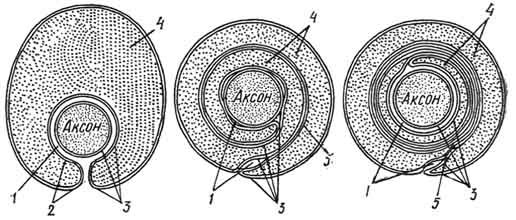
Рис. 163. Схема развития миелинового волокна:
1 — контакт аксолеммы и оболочки леммоцита; 2 — щель; 3 — аксолемма и оболочки леммоцита; 4 — цитоплазма леммоцита; 5 — мезаксон.
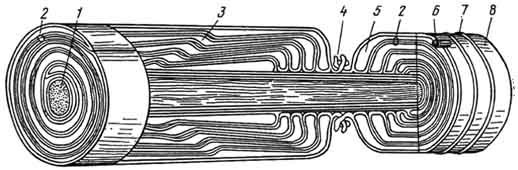
Рис. 164. Схема строения миелинового волокна:
1 — аксон; 2 — мезаксон; 3 — насечки; 4 — перехват; 5 — протоплазма леммоцита; 6 — ядро леммоцита; 7 — неврилемма; S — эндоневрий.
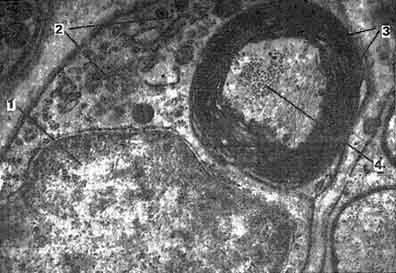
Рис. 165. Электронная микрофотография поперечного среза миелинового волокна из краниального шейного узла крупного рогатого скота (препарат Козлова):
1 — ядро леммоцита; 2 — цитоплазма леммоцита; 3 — пластины миелиновой оболочки; 4 — нейрофиламенты в цитоплазме аксона.
Осевой цилиндр нервных волокон состоит из нейроплазмы — цитоплазмы нервных клеток, содержащей продольно ориентированные нейрофиламенты и нейротубулы. Наличие различных органелл и их локализация специфичны для осевых цилиндров волокон различного функционального значения.