
Различают два осн. типа Р. Всем прокариотич. организмам (бактерии и синезеленые водоросли) свойственны т. наз. 70S Р., характеризующиеся коэф. (константой) седиментации ок. 70 единиц Сведберга, или 70S (по коэф. седиментации различают и Р. др. типов, а также субчастицы и биополимеры, входящие в состав Р.).
Их мол. м. составляет 2,5
- 10 6 , линейные размеры 20-25 нм. По хим. составу это рибонуклеопротеиды;
- они состоят только из рРНК и белка (соотношение этих компонентов 2:1).
Рибо-сомная РНК в Р. присутствует гл. обр. в виде Mg-соли (по-видимому, частично и в виде Са-соли);
- магния в Р. до 2% от сухой массы. Кроме того, в разл. кол-вах (до 2,5%) могут присутствовать также катионы аминов-спермина H 2 N(CH 2 ) 3 NH(CH 2 ) 4 NH(CH 2 ) 3 NH 2 , спермидина H 2 N(CH 2 ) 3 NH(CH 2 ) 4 NH 2 и др.
Цитоплазма клеток всех эукариотич. организмов содержит неск. более крупные 80S Р. Их мол. м. ок. 4·10 6 , линейные размеры 25-30 нм, содержание белка в них значительно больше, чем в прокариотической Р. (соотношение РНК: белок ок. 1:1).
Рибосомная РНК 80S также связана в осн. с Mg и Са и с небольшим кол-вом полиаминов (спермин, спермидин и др.).
Хлоропласты и митохондрии эукариотич. клеток содержат Р., отличные от типа 80S. Р. хлоропластов высших растений принадлежат к истинному 70S типу. Митохонд-риальные Р. более разнообразны; их строение находится в зависимости от таксономич. принадлежности организма (т.е. от принадлежности к определенному виду, роду или семейству).
Напр., митохондриальные Р. млекопитающих существенно мельче типичных 70S Р.; коэф. седиментации этих Р. составляет ок. 55S (т. наз. минирибосомы).
Р. из самых разнообразных организмов (как прокариотич., так и эукариотич.) имеют сходное строение. Они состоят из двух разделяемых субчастиц, или рибосомных субъединиц. При определенных условиях (напр., при понижении концентрации Mg 2 + в среде) Р. обратимо диссоциирует на две субчастицы с соотношением их мол. масс ок. 2:1. Прокарйотическая 70S Р. диссоциирует на субъединицы с коэф. седиментации 50S (мол. м. 1,5·10 6 ) и 30S (мол. м. 0,85·10 6 ).
Эукариотическая Р. разделяется на субчастицы 60S и 40S. Две рибосомные субчастицы объединены в полную Р. строго определенным образом, предполагающим специфич. контакты их поверхностей.
Подземный ремонт скважин. Виды подземных ремонтов, применяемая ...
... и эксплуатацию производят без крепления ствола скважины. Устье скважины в зависимости ... рых все стволы выстреливают одновременно — залпом. Такие перфораторы ... 3. Перфораторы селективного, или раздельного, выборочного действия, дающие ... Этот участок скважины закрепляют обсадной трубой 1 (состоящей из свинченных ... ПЕРФОРИРОВАНИЕ ТРУБ В СКВАЖИНАХ. Конструкция скважины в виде сплошной эксплуатационной ...
Как прокариотические, так и эукариотические Р. содержат две разл. высокомол. рРНК (по одной на каждую субчастицу) и одну относительно низкомол. рРНК в большой субчастице.
Рибосомные белки большинства животных представлены в осн. умеренно основными полипептидами, хотя имеется неск. нейтральных и кислых белков. Мол. м. рибосомных белков варьирует от 6 тыс. до 60 тыс. В прокариотической Р. малая субчастица (30S) содержит ок. 20, большая (508)-ок. 30 разл. белков; в эукариотической P. 40S субчастица включает ок. 30 белков, а 60S-ок. 40 (обычно Р. не содержат двух или неск. одинаковых белков).
Рибосомные белки характеризуются глобулярной компактной конформацией с развитой вторичной и третичной структурой; они занимают преим. периферич. положение в ядре, состоящем из рРНК. В отличие от вирусных нуклеопротеидов в структурно асим. рибосомном нуклеопротеиде рРНК не покрыта сплошной белковой оболочкой, а в ряде мест образует пов-сть Р. Плотность упаковки рРНК в Р. достаточно высока и приблизительно соответствует плотности кристаллич. упаковки гидратир. полинуклеотидов.
По-видимому, рРНК определяет осн. структурные и функцион. св-ва Р, в частности обеспечивает целостность рибосомных субъединиц, обусловливает их форму и ряд структурных особенностей. Специфич. пространств. структура рРНК детерминирует локализацию всех рибосомных белков, играет ведущую роль в организации функцион. центров Р.
Рибосомный синтез белка-многоэтапный процесс. Первая стадия (инициация) начинается с присоединения матричной РНК (мРНК) к малой рибосомной субчастице, не связанной с большой субчастицей. Характерно, что для начала процесса необходима именно диссоциированная Р. К образовавшемуся т. наз. инициаторному комплексу присоединяется большая рибосомная субчастица. В стадии инициации участвуют спец. инициирующий кодон (см. Генетический код), инициаторная транспортная РНК (тРНК) и специфич. белки (т. наз. факторы инициации).
Пройдя стадию инициации, Р. переходит к последоват. считыванию кодонов мРНК по направлению от 5′- к 3′-концу, что сопровождается синтезом полипептидной цепи белка, кодируемого этой мРНК (подробнее о механизме синтеза полипептидов см. в ст. Трансляция).
В этом процессе Р. функционирует как циклически работающая мол. машина. Рабочий цикл Р. при элонгации состоит из трех тактов: 1) кодонзави-симого связывания аминоацил-тРНК (поставляет аминокислоты в Р.), 2) транспептидации-переноса С-конца растущего пептида на аминоацил-тРНК, т.е. удлинения строящейся белковой цепи на одно звено, 3) транслокации-перемещения матрицы (мРНК) и пептидил-тРНК относительно Р. и переход Р. в исходное состояние, когда она может воспринять след. аминоацил-тРНК. Когда Р. достигнет специального терминирующего кодона мРНК, синтез полипептида прекращается. При участии специфич. белков (т. наз. факторов терминации) синтезир. полипептид освобождается из Р. После терминации Р. может повторить весь цикл с др. цепью мРНК или др. кодирующей последовательностью той же цепи.
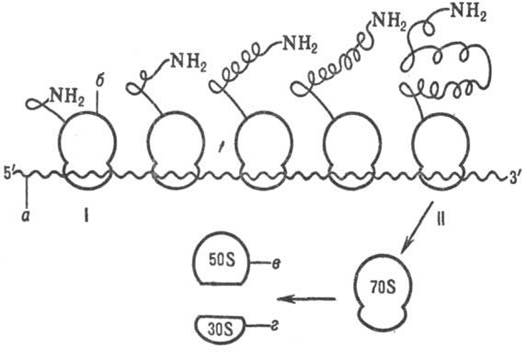
Схема синтеза полипептидной цепи полирибосомой: I-начал о синтеза, II-окончание синтеза; а-мРНК, б-рибосома, в-большая субъединица рибосомы, г-малая субъединица рибосомы.
Метаболизм как основа жизнедеятельности клетки
... соответствии с генетической программой клетки, реализуя ее наследственную информацию. Энергетический обмен в клетке. Синтез АТФ Человек и ... katabole разрушение), или энергетического обмена. Примерами таких реакций является расщепление липидов, полисахаридов, белков и нуклеиновых кислот ... с образованием токсичного аммиака. Белки и составляющие их аминокислоты используются клеткой для получения энергии ...
В клетках с интенсивной секрецией белка и развитым эндоплазматич. ретикулумом значит. часть цитоплазмати-ческой Р. прикреплена к его мембране на пов-сти, обращенной к цитоплазме. Эти Р. синтезируют полипептиды, к-рые непосредственно транспортируются через мембрану для дальнейшей секреции. Синтез полипептидов для внутриклеточных нужд происходит в осн. на свободных (не связанных с мембраной) Р. цитоплазмы. При этом транслирующие Р. не равномерно диспергированы в цитоплазме, а собраны в группы. Такие агрегаты Р. представляют собой структуры, где мРНК ассоциирована со многими Р., находящимися в процессе трансляции; эти структуры получили назв. полирибосом или полисом.
При интенсивном синтезе белка расстояние между Р. вдоль цепи мРНК в полирибосоме м. б. предельно коротким, т.е. Р. находятся почти вплотную друг к другу. Р., входящие в полирибосомы, работают независимо и каждая из них синтезирует полную полипептидную цепь (см. рис.).
Р ибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК).
Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма —с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока).
Рибосомы. Так же как и эндоплазматическая сеть, рибосомы были открыты только с помощью электронного микроскопа. Рибосомы — самые маленькие из клеточных органелл.
Рибосомы либо располагаются на поверхности мембраны гранулярной ЭПС в один ряд, либо образуют розетки и спирали. В тех клетках, где хорошо развита гранулярная ЭПС, например, в полностью дифференцированных клетках печени и поджелудочной железы, большинство рибосом связано с ее мембранами. В клетках же, где гранулярная ЭПС развита слабо, рибосомы преимущественно свободно располагаются в основном веществе цитоплазмы. К клеткам такого типа относятся плазмоциты лимфатических узлов и селезенки, овоциты человека и ряд других. Помимо цитоплазмы, рибосомы обнаружены и в клеточном ядре, где они имеют такую же округлую форму, строение и размеры, как и рибосомы цитоплазмы. Часть ядерных рибосом свободно располагается в кариоплазме, а часть их находится в связи с нитевидными структурами, из которых состоят остаточные хромосомы, обнаруживаемые обычно при электронномикроскопическом исследовании интерфазного ядра. В последнее время рибосомы обнаружены в митохондриях и пластидах клеток растений.
Биохимический анализ рибосом, полученных путем дифференциального центрифугирования клеточных гомогенатов, показал, что в состав их входит высокополимерная, так называемая рибосомальная РНК и белок. Соотношение этих двух компонентов в рибосомах почти одинаково.
Строение и функции клетки
... В цитоплазме расположены мельчайшие структуры – органоиды. К органоидам клетки относятся: эндоплазматическая сеть, рибосомы, митохондрии, лизосомы, комплекс Гольджи, клеточный центр. Мембрана. Если рассматривать в микроскоп клетку ... и в 1898 году был назван «комплексом (аппаратом) Гольджи». Белки, выработанные в рибосомах, поступают в комплекс Гольджи, а когда они требуются другому органоиду, ...
Белок рибосом самых разнообразных клеток и разных организмов в общем одинаков по составу аминокислот, причем в нем часто преобладают основные аминокислоты, а следовательно, белки рибосом имеют …? свойства. Рибосомы содержат также Mg2+.
Функции рибосом. Исследование ультраструктуры клеток многочисленных видов многоклеточных растений и животных, бактерий и простейших показало, что рибосомы — обязательный органоид каждой клетки. Наличие этого органоида во всех клетках, однородность его строения и химического состава свидетельствуют о важной роли рибосом в жизнедеятельности клеток. Было выяснено, что на рибосомах происходит синтез белков.
В процессах биосинтеза белка роль рибосом заключается в том, что к ним из основного вещества цитоплазмы непрерывно подносятся с помощью т-РНК аминокислоты, и происходит укладка этих аминокислот в полипептидные цепи в строгом соответствии с той генетической информацией, которая передается из ядра в цитоплазму через и-РНК, постоянно поступающую к рибосомам. На основании такой функции рибосом в белковом синтезе можно назвать их своего рода «сборочными конвейерами», на которых в клетках образуются белковые молекулы.
В процессе синтеза белка, таким образом, активное участие принимают т-РНК и и-РНК, а роль рибосомальной РНК еще не выяснена. По имеющимся в настоящее время данным, рибосомальная РНК не принимает участия в синтезе белковых молекул. В комплексе с белком рибосом она образует строму этого органоида.
При осуществлении процессов синтеза белка в клетках активную роль выполняют не все рибосомы. Специальные биохимические исследования позволили установить. Что наиболее активная роль в синтезе клеточных белков принадлежит рибосомам, связанным с мембранами ЭПС. Можно предполагать, что эти два органоида, теснейшим образом связанные друг с другом, представляют собой единый аппарат синтеза (рибосомы) и транспорта (эндоплазматическая сеть) основной массы белка, вырабатываемого в клетке.
В рибосомах. Находящихся в ядре, происходит синтез ядерных белков. Рибосомы митохондрий и пластид выполняют функцию синтеза части белков, содержащихся в этих органоидах.
Вопрос о том, где в клетке образуются рибосомы, до сих пор не решен, но сейчас уже довольно убедительно показано, что основным местом формирования рибосом служит ядрышко и образованные в нем рибосомы поступают из ядра в цитоплазму.
1. История исследования рибосомы
Рибосомы впервые были описаны как уплотненные частицы, или гранулы, клеточным биологом румынского происхождения Джорджем Паладе в середине 1950-х годов. В 1974 г. Паладе, Клод и Кристиан Де Дюв получили Нобелевскую премию по физиологии и медицине «за открытия, касающиеся структурной и функциональной организации клетки». Термин «рибосома» был предложен Ричардом Робертсом в 1958 вместо «рибонуклеобелковая частица микросомальной фракции». Биохимические и мутационные исследования рибосомы начиная с 1960-х позволили описать многие функциональные и структурные особенности рибосомы. В начале 2000-х появились атомные структуры отдельных субъединиц, а также полной рибосомы, связанной с различными субстратами, которые позволили понять механизм декодинга (распознавания антикодона тРНК, комплементарного кодону мРНК) и детали взаимодействий между рибосомой, антибиотиками, тРНК и мРНК.
Производство кормовых белков
... Ввиду актуальности исследования и разработки технологий микробиологического производства кормовых белковых препаратов целью данной работы является проектирование линии производства дрожжевого белка. 1. Химический состав кормовых дрожжей Биомасса дрожжевых клеток состоит из ...
2. Строение и функции, разновидности рибосом
Рибосома — крупный внутриклеточный макромолекулярный ансамбль, ответственный за синтез полипептидной цепи из аминокислот (трансляцию); состоит из молекул РНК (т. наз. рибосомные рибонуклеиновые кислоты, или рРНК) и белков.
Основная масса рибосом локализована в цитоплазме. В бактериальной клетке рибосомы составляют до 30% ее сухой массы: на одну бактериальную клетку приходится примерно 104 рибосом. В эукариотических клетках (клетки всех организмов, за исключением бактерий и синезеленых водорослей) относительное содержание рибосом меньше, и их количество очень сильно варьирует в зависимости от белок-синтезирующей активности соответствующей ткани или отдельной клетки.
В эукариотической клетке все рибосомы цитоплазмы (как мембрано-связанные, так и свободные) образуются в ядрышке; считается, что там они неактивны. Эукариотическая клетка имеет также специальные рибосомы в митохондриях (у животных и растений) и хлоропластах (у растений).
Рибосомы этих органелл отличаются от цитоплазматических размерами и некоторыми функциональными свойствами. Они образуются непосредственно в этих органеллах.
Различают два основных типа рибосом. Всем прокариотическим организмам (бактерии и синезеленые водоросли) свойственны так называемые 70 S рибосомы, характеризующиеся коэффициентом (константой) седиментации около 70 единиц Сведберга, или 70 S (по коэф. седиментации различают и рибосомы других типов, а также субчастицы и биополимеры, входящие в состав Р.).
Их молекулярная масса составляет 2,5
- 10 6 , линейные размеры 20-25 нм. По химическому составу это рибонуклеопротеиды;
- они состоят только из рРНК и белка (соотношение этих компонентов 2:1).
Рибосомная РНК в рибосоме присутствует в основном в виде Mg -соли (по-видимому, частично и в виде Са-соли);
- магния в рибосоме до 2% от сухой массы. Кроме того, в различных количествах (до 2,5%) могут присутствовать также катионы аминов-спермина H 2 N ( CH 2)3 NH ( CH 2)4 NH ( CH 2)3 NH 2, спермидина H 2 N ( CH 2)3 NH ( CH 2)4 NH 2 и др.
Поскольку коэффициенты седиментации зависят не только от молекулярной массы, но и от формы частиц, седиментационные коэффициенты при диссоциации неаддитивны: так, например, бактериальные рибосомы с молекулярной массой ~3*106 Дальтон имеет коэффициент седиментации 70S, обозначается как 70S и диссоциирует на субъединицы 50S и 30S: 70S 50S + 30S
Рибосомные субчастицы содержат по одной молекуле рРНК большой длины, масса которой составляет ~1/2 — 2/3 массы рибосомной субчастицы, таким образом, в случае бактериальных рибосом 70S субчастица 50S содержит рРНК 23S (длина ~3000 нуклеотидов) и субчастица 30S содержит рРНК 16S (длина ~1500 нуклеотидов); большая рибосомная субчастица кроме «длинной» рРНК содержит также одну или две «коротких» рРНК (5S рРНК бактериальных рибосомных субчастиц 50S или 5S и 5.8S рРНК больших рибосомных субчастиц эукариот).
Митохондрии как энергетические станции клеток
... 22 различных тРНК. Имея собственный генетический аппарат, митохондрия обладает и собственной белоксинтезирующей системой, особенностью которой в клетках животных и грибов являются очень маленькие рибосомы, характеризуемые ... собой пространство между наружной и внутренней мембранами митохондрии. Его толщина — 10-20 нм. Так как наружная мембрана митохондрии проницаема для небольших молекул и ионов, ...
Цитоплазма клеток всех эукариотических организмов содержит несколько более крупные 80 S рибосомы. Их молекулярная масса около 4·10 6 , линейные размеры 25-30 нм, содержание белка в них значительно больше, чем в прокариотической рибосоме (соотношение РНК: белок ок. 1:1).
Рибосомная РНК 80 S также связана в основном с Mg и Са и с небольшим кол-вом полиаминов (спермин, спермидин и др.).
Хлоропласты и митохондрии эукариотических клеток содержат рибосомы, отличные от типа 80 S . Рибосомы хлоропластов высших растений принадлежат к истинному 70 S типу. Митохондриальные рибосомы более разнообразны; их строение находится в зависимости от таксономической принадлежности организма (т.е. от принадлежности к определенному виду, роду или семейству).
Напр., митохондриальные рибосомы млекопитающих существенно мельче типичных 70 S Р.; коэффициент седиментации этих рибосом составляет около 55 S (т. наз. минирибосомы).
Рибосомы из самых разнообразных организмов имеют сходное строение. Они состоят из двух разделяемых субчастиц, или рибосомных субъединиц. При определенных условиях (например, при понижении концентрации Mg 2 + в среде) рибосома обратимо диссоциирует на две субчастицы с соотношением их молекулярных масс около 2:1. Прокарйотическая 70 S рибосома диссоциирует на субъединицы с коэффициентом седиментации 50 S (молекулярная масса 1,5·10 6 ) и 30 S (молекулярная масса 0,85·10 6 ).
Эукариотическая рибосома разделяется на субчастицы 60 S и 40 S . Две рибосомные субчастицы объединены в полную рибосому строго определенным образом, предполагающим специфические контакты их поверхностей.
Как прокариотические, так и эукариотические рибосомы содержат две различные высокомолекулярные рРНК (по одной на каждую субчастицу) и одну относительно низкомолекулярную рРНК в большой субчастице.
Рибосомные белки большинства животных представлены в основном умеренно основными полипептидами, хотя имеется несколько нейтральных и кислых белков. Молекулярные массы рибосомных белков варьирует от 6 тыс. до 60 тыс. г/моль. В прокариотической рибосоме малая субчастица (30 S ) содержит около 20, большая (50 S ) — около 30 различных белков; в эукариотической рибосоме 40 S субчастица включает около 30 белков, а 60 S -около 40 (обычно рибосомы не содержат двух или нескольких одинаковых белков).
Рибосомные белки характеризуются глобулярной компактной конформацией с развитой вторичной и третичной структурой; они занимают преимущественно периферическое положение в ядре, состоящем из рРНК. Плотность упаковки рРНК в рибосомах достаточно высока.
По-видимому, рРНК определяет основные структурные и функциональные свойства рибосом, в частности обеспечивает целостность рибосомных субъединиц, обусловливает их форму и ряд структурных особенностей. Специфическая пространственная структура рРНК детерминирует локализацию всех рибосомных белков, играет ведущую роль в организации функциональных центров рибосом.
Разделение декодирующей и энзиматической функций между субчастицами
Трансляция начинается с того, что мРНК, синтезируемая на ДНК в качестве копии одной из двух цепей последней, связывается с рибосомной частицей. При этом рибосомная частица (у прокариот прямо и непосредственно, а у эукариот после некоторого скольжения вдоль некодирующей части мРНК) специфически взаимодействует с началом кодирующей нуклеотидной последовательности мРНК. Этап связывания мРНК с рибосомной частицей и нескольких последующих событий, приводящих к образованию первой пептидной связи, называется инициацией трансляции. Вслед за инициацией рибосома последовательно «читает» цепочку мРНК по тройкам (триплетам) нуклеотидов по направлению к 3′-концу, наращивая (удлиняя) полипептидную цепочку аминокислотными остатками; этот этап собственно трансляции называется элонгацией . Наконец, достигнув специального нуклеотидного триплета — стоп-кодона, или кодона терминации, — рибосома освобождает синтезированную полипептидную цепочку белка: происходит терминация трансляции.
Нарушения белкового обмена
... использоваться как энергетический материал. Таким образом, белковый обмен ... мРНК к рибосомам, а также процессу транслокации могут тормозиться дифтерийным токсином. К ингибиторам инициации можно отнести и ряд широко используемых антибактериальных антибиотиков, тормозящих присоединение аминоацил–тРНК ... заболеваний, характеризующихся наследственной недостаточностью каталитической активности отдельных ...
Генетические функции малой рибосомной субчастицы.
Характерным моментом инициации трансляции является то, что на этом этапе участвуют не целые рибосомы, а их отдельные субчастицы. Другими словами, для того чтобы инициировать трансляцию, рибосома должна быть диссоциирована на две составляющие ее неравные субчастицы. Для этого клетка располагает специальными механизмами, обеспечивающими диссоциацию рибосом после терминации трансляции. Именно малая субчастица рибосомы (30S у прокариот и 40S у эукариот), и только она, связывается с мРНК, то есть служит первичным приемником генетической информации для белоксинтезирующего аппарата. Лишь впоследствии, при завершении этапа инициации трансляции, к ней присоединяется большая субчастица (50S у прокариот и 60S у эукариот), образуя полную рибосомную частицу (70S у прокариот и 80S у эукариот), которая и будет производить элонгацию.
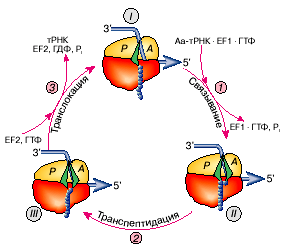
Рис. 1: Элементарный элонгационный цикл рибосомы, в результате которого прочитывается один триплет (кодон) мРНК и образуется одна пептидная связь (добавляется одна аминокислота к растущему полипептиду).
В процессе элонгации рибосома удерживает мРНК и движется относительно ее (или протягивает ее сквозь себя) в направлении от 5′-конца к 3′-концу. Удержание мРНК на рибосоме есть целиком и полностью функция малой рибосомной субчастицы, в то время как большая субчастица с мРНК никак не взаимодействует. Соответственно последовательное сканирование кодирующей последовательности мРНК (считывание генетической информации) в ходе элонгации осуществляется на малой субчастице транслирующей рибосомы.
Механизм потриплетного сканирования мРНК в ходе элонгации предполагает участие молекул тРНК, которые взаимодействуют прежде всего с малой рибосомной субчастицей. Малая субчастица в составе полной транслирующей рибосомы имеет два тРНК-связывающих участка, обозначаемых как аминоацил-тРНК-связывающий участок (А-участок) и пептидил-тРНК-связывающий участок (Р-участок).
На этапе элонгации Р-участок всегда занят остатком тРНК.
Рассмотрение элементарного акта элонгации удобно начать с момента, когда Р-участок занят молекулой пептидил-тРНК (тРНК, несущая растущую полипептидную цепь), а А-участок вакантен и содержит лишь некий нуклеотидный триплет (кодон) мРНК, пока не взаимодействующий ни с каким триплетом (антикодоном) тРНК (рис. 1, состояние I).
Технология рекомбинантных ДНК
... исследователей, дали очень обнадеживающие результаты. дезоксирибонуклеиновый ген секвенирование 1. Конструирование рекомбинантных ДНК Под рекомбинантными понимают ДНК, образованные объединением in vitro (в пробирке) двух или более ... какой-либо регуляторный генетический элемент, например, промотор или участок, связанный с рибосомой. В этом случае линкеры обеспечивают не только объединение генов, но ...
Такая рибосома готова (компетентна) связать аминоацил-тРНК, антикодон которой комплементарен триплету (кодону), установленному в А-участке. При наличии около рибосомы такой аминоацил-тРНК происходит первый шаг элементарного элонгационного цикла — кодонспецифическое связывание аминоацил-тРНК с А-участком. Теперь рибосома несет «старую» пептидил-тРНК в Р-участке и новоявленную аминоацил-тРНК в А-участке, которые расположены рядом, бок о бок (рис. 1, состояние II ).
Следовательно, в результате кодон-антикодонового взаимодействия мРНК с тРНК на малой субчастице рибосомы произошло декодирование триплета мРНК: именно тот аминокислотный остаток, который был привешен к тРНК с комплементарным антикодоном, оказался в рибосоме.
Далее молекулы пептидил-тРНК и аминоацил-тРНК, расположенные рядом в рибосоме, реагируют друг с другом: пептидильный остаток переносится на аминогруппу молекулы аминоацил-тРНК. Это второй шаг элементарного элонгационного цикла — транспептидация, когда полипептидная цепь удлиняется на одну аминокислоту — на ту, которую принесла тРНК, связавшаяся с А-участком. А сама тРНК, принесшая эту аминокислоту, так и осталась с ней связанной и, таким образом, связанной с удлиненным полипептидом (рис. 1, состояние III).
В этом состоянии, однако, новообразованная пептидил-тРНК — точнее, ее остаток тРНК — занимает «не положенный ей» А-участок, а «сидит» в Р-участке деацилированная (без пептидильного или аминоацильного остатков) тРНК. Такое состояние называется претранслокационным. Дальше элонгация идти не может, пока не осуществится третий шаг элонгационного цикла — транслокация, которая выбросит деацилированную тРНК из Р-участка и переведет пептидил-тРНК из А-участка в Р-участок вместе со связанным с ней кодоном мРНК. В результате в освободившемся А-участке на малой рибосомной субчастице установится следующий (новый) кодон мРНК.
Цикл завершился, приведя к образованию одной пептидной связи и соответствующему удлинению растущего полипептида на одну аминокислоту, с одной стороны, и к прочтению одного кодона мРНК и перемещению мРНК на один триплет — с другой. Повторение таких элементарных циклов и создает процесс элонгации.
Таким образом, малая рибосомная субчастица в изолированном состоянии воспринимает копию гена в форме мРНК и инициирует процесс ее трансляции, а в ходе трансляции малая субчастица полной рибосомы удерживает мРНК на себе, декодирует ее с помощью тРНК и последовательно перебирает ее кодоны и тРНК, используя механизм транслокации. Так как все это операции с генетическим материалом, то указанные функции малой рибосомной субчастицы могут быть определены как генетические.
рибосома биосинтез белок
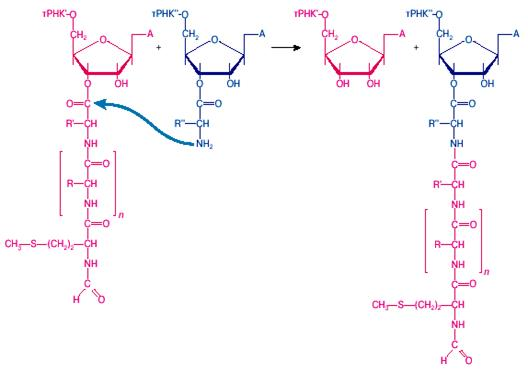
Рис. 2: Реакция транспептидации, катализируемая пептидилтрансферазным центром большой рибосомной субчастицы
Энзиматические функции большой рибосомной субчастицы.
Когда пептидил-тРНК занимает Р-участок, а аминоацил-тРНК — А-участок на малой субчастице рибосомы (см. рис. 1, состояние II) , концы остатков тРНК с присоединенными к ним аминоацильными остатками взаимодействуют с большой субчастицей рибосомы. Участок этого взаимодействия на большой субчастице является пептидилтрансферазным центром рибосомы: он катализирует реакцию транспептидации между пептидил-тРНК и аминоацил-тРНК, то есть перенос карбоксильной группы пептидильного остатка на аминогруппу аминоацил-тРНК (рис. 2).
Основы гистологической техники. Цитохимические методы
... заключают в водорастворимую среду(глицерин, желатин и др.). Этапы изготовления гистологических препаратов. При изготовлении препаратов выполняют следующие операции. 1. Фиксируют ... приблизительная качественная оценка, основанная на степени интенсивности окраски. мейоз плазмалемма гистологический цитоплазматический 2. Наружная цитоплазматическая мембрана (плазмалемма). Её строение и функция. ...
В результате образуется новая пептидная связь, и пептидильный остаток становится на одну аминокислоту длиннее. Таким образом, большая субчастица транслирующей рибосомы выступает здесь как фермент, ответственный за образование пептидных связей и в целом за синтез (элонгацию) полипептидной цепи. Это главная энзиматическая функция рибосомы.
Следует отметить, что никакого отдельного от рибосомы белка-фермента, катализирующего образование пептидных связей на рибосоме, не существует. Не найдено и никакого специального белка в составе рибосомы, который бы обладал такой энзиматической функцией. Транспептидация катализируется пептидилтрансферазным центром самой рибосомы как интегральной частью большой рибосомной субчастицы, и основной вклад в организацию центра вносит, по-видимому, рибосомная РНК субчастицы.
Кроме катализа реакции транспептидации большая рибосомная субчастица определенным образом участвует в энзиматическом расщеплении (гидролизе) гаунозинтрифосфата (ГТФ) в процессе трансляции. Дело в том, что, как видно на рис. 1, первый и третий шаги элонгационного цикла идут с участием специальных нерибосомных белков — так называемых факторов элонгации EF1 и EF2. Эти белки являются катализаторами соответствующих нековалентных переходов — связывания аминоацил-тРНК и транслокации. Для такого катализа необходимым оказывается сопряженный гидролиз ГТФ. Именно большая рибосомная субчастица взаимодействует с факторами элонгации и индуцирует гидролиз ГТФ на них. Хотя сам ГТФазный центр находится не на рибосомной субчастице, а на белке — факторе элогнации, ее временная ассоциация с фактором существенна для формирования активного энзиматического ГТФазного центра.
Таким образом, существует четкое разделение труда между двумя неравными субчастицами рибосомы: малая субчастица выполняет генетические функции, будучи ответственной за прием и декодирование генетической информации, в то время как большая участвует в энзиматических реакциях в процессе трансляции.
Конформационная подвижность рибосомы
Работа рибосомы в качестве «лентопротяжного механизма» последовательное прочитывание цепи мРНК от одного конца к другому) в ходе элонгации и ее способность перебрасывать сравнительно большие молекулярные массы (молекулы тРНК) из одного участка в другой в каждом элементарном элонгационном цикле (см. рис. 1, шаг 3 ) предполагают ее механическую подвижность. Взаимная подвижность двух рибосомных субчастиц может быть основным видом крупноблочной подвижности рибосомы в ходе работы, и имеются экспериментальные свидетельства в пользу такой подвижности. Кроме того, существуют указания на подвижность «головки» малой рибосомной субчастицы относительно ее «тела» и на подвижность палочкообразного бокового выступа большой рибосомной субчастицы.
Рибосома при прохождении элонгационного цикла осциллирует между двумя конформационными состояниями: закрытым (сомкнутым) и открытым (разомкнутым).
В сомкнутом состоянии рибосомные лиганды (тРНК) зажаты между субчастицами, связаны максимальным количеством контактов с рибосомой и не имеют внутририбосомной подвижности. В разомкнутом состоянии рибосомы лиганды более подвижны, контакты с рибосомой менее полны, и имеется возможность их входа и выхода из рибосомы. Так, на первом этапе связывания аминоацил-тРНК рибосома должна быть открыта для приема лиганда. Возможно, это открытое состояние фиксируется фактором элонгации EF1. Далее EF1 уходит, рибосомные субчастицы плотно смыкаются, и аминоацильный конец связавшейся аминоацил-тРНК вступает в контакт с пептидилтрансферазным центром большой субчастицы. В сомкнутом состоянии пептидил-тРНК и аминоацил-тРНК тесно сближены, и между ними происходит реакция транспептидации. Теперь, чтобы выбросить деацилированную тРНК из рибосомы и дать свободу для перемещения остатка тРНК молекулы пептидил-тРНК из А-участка в Р-участок, рибосому надо приоткрыть, в частности путем раздвигания субчастиц. Это может осуществляться фактором элонгации EF2. После ухода EF2 с рибосомы она снова смыкается и ждет прихода очередной аминоацил-тРНК с фактором элонгации EF1.
Процесс периодического смыкания-размыкания рибосомы является энергозависимым: факторы элонгации EF1 и EF2 взаимодействуют с рибосомой только будучи связанными с ГТФ (согласно модели, при этом взаимодействии происходит открывание рибосомы), а взаимодействие с рибосомой наводит ГТФазную активность, ГТФ гидролизуется, фактор элогации теряет сродство к рибосоме и уходит, и рибосома закрывается. Таким образом, на каждое смыкание-размыкание рибосомы расходуется одна молекула ГТФ. Так как в каждом элонгационном цикле рибосома смыкается-размыкается дважды, то две молекулы ГТФ расходуются на каждый цикл. Это есть энергетическая плата за эффективное (быстрое и надежное) функционирование рибосомы как молекулярной машины.
4. Список литературы
[Электронный ресурс]//URL: https://inzhpro.ru/referat/ribosomyi-fabrika-sinteza-belka/
1. Спирин А.С. Принципы структуры рибосом // Соросовский Образовательный Журнал. 1998. N 11. С. 65-70.
2. Спирин А.С. Молекулярная биология: Структура рибосомы и биосинтез белка. М.: Высш. шк., 1986. 300 с.
3. Спирин А.С. О механизме работы рибосомы: Гипотеза смыкания-размыкания субчастиц // Докл. АН СССР. 1968. Т. 179. С. 1467-1470.